ミーハースポーツ放浪記
大野さんに,分泌にまつわるエッセイを,と依頼され,はじめ「分泌研究30年」というタイトルをつけてみたのだが,これはつくづく自分の年を感じさせるものであるとすぐにやめにした。とは言っても,書こうとしているのはそんなような内容である。正確に言うと,私がこの世界に入ったのは1980年だから,自分の歴史としてはまだ30年は経っていない。いま大学で細胞生物学を教えていて,分泌の話は1960年代の膜結合型ポリソームの発見あたりから入ることにしているのだが,分子レベルの研究に向かい始めるのは1972年のシグナルペプチドの発見1),そして1975年の金字塔「シグナル仮説」の提唱2)が大きな契機である。そこらへんから数えるとやはり30年くらいの歴史になる。そのころ学生だった私がこの分野の黎明を知り,次第に足を踏み入れていったのにはいろいろ偶然も重なっているのだが,ほとんど全てが未知であったおもしろい時代にこの世界に入れて幸運だったと思っている。自分自身のことを振り返りながら,少し雑感など綴ってみることにする。
●ロックフェラーグループの独壇場時代
1970年代の分泌研究は,George Palade(1974年ノーベル賞),David Sabatini,Günter Blobel(1999年ノーベル賞)といった,当時のロックフェラー大学のグループの独壇場だった3)。わが国でも,大村恒雄さん,田代裕さん,そして少し遅れて三原勝芳さんがロックフェラー大学の留学から帰られて,この領域で指導的な役割を果たされることになる。学生だった私はまだこの業界には入っておらず,博士課程時代に出入りさせてもらっていた東大医科研の上代研究室(当時タンパク質生合成の研究のメッカであった)の論文紹介で聞いて,「分泌の信号,へーそんなもんがあるんだ」と思う程度だった。私のあまり知られていない過去なのだが,そのころはNMRが専門で,EF-Tuの構造解析をしていた。
それが,1980年に学位を取って国立予防衛生研究所(予研,今の国立感染症研究所),赤松穰部長率いる化学部に研究員として就職し,脂質の観点から見た生体膜の研究をせよ,と命ぜられて人生が変わる。今までタンパク合成をやってきたのだから,分泌におけるリン脂質の役割を調べてはどうかというアドバイスを受け,勉強を始めてあっという間にはまり込んだ。ちょうど小胞体膜透過の分子装置が見つかりかけたころであり,SRP(signal recognition particle; はじめはsignal recognition proteinだった)およびSRP受容体(はじめはdocking proteinと呼ばれていた)の発見など,David Meyer,Bernhard DobbersteinのEMBLグループと,Peter Walter,G. Blobelのロックフェラーグループが大バトルを繰り広げようとしている時期でもあった。
予研の研究室では動物細胞が材料であり,まずは培養細胞の取り扱い,ウイルスの取り扱い,脂質の生化学などを習った。1年先輩であった前田正知さんが,VSV(水疱性口内炎ウイルス)を用いて細胞内輸送を調べる系を作り始めていたが,免疫学的な手法など独力で学ばなくてはならないことも多かった。そもそもVSVも日本では規制が厳しく,非常に神経を使いながらの実験だった。私が動物細胞の研究をしていたということは,あまり知られていない過去その2である。
●カリフォルニアグループの台頭
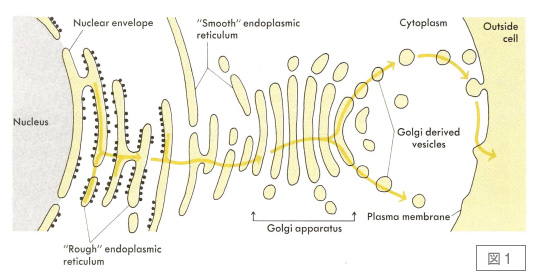
マウスミエローマ細胞を使って,リン脂質組成を人工的に変えてやると,VSVのウイルス産生や,VSV Gタンパク質の細胞内輸送,免疫グロブリンの分泌,細胞内輸送が遅くなってくる。はじめは,小胞体の膜透過に差が出ているんだろうと思っていたが,次第に,これは小胞体以降の過程に影響があるんじゃないかと思い始めた。まだ小胞輸送という言葉も定着しておらず,小胞体,ゴルジ体から細胞外まで,だらだらとつながっているような絵がしばしば書かれ(図1),小胞体の膜さえ越えてしまえばあとは勝手に細胞表面,細胞外まで流れていってしまう,というような考え方が普通であって,オルガネラ間の輸送に何らかの分子装置がいるのかどうか,という今なら信じられないような議論が堂々と行われていた時代である。このへんの問題がこれから重要になっていくのではないかと思い,よし小胞体以降の輸送を自分の研究テーマにしよう,と心に決めたのはこのころのことであった。
ここで私は,とても恥ずかしい秘密を1つ告白する。実は,NovickとSchekmanによる酵母sec 変異株分離の最初の論文(PNAS, 1979)4)を見過ごしていたのである。1980年にCellに続報5)が掲載されて,慌ててまとめて読んだが,その本当のすごさがよくわかったのは,自分自身でCHO細胞の分泌変異株をスクリーニングしようと思い立ってからのことであった。
私自身が留学することになった先のボスであり,またその後は最強の競争相手にもなって,いろんな意味で私にとても大きな影響を与えたRandy Schekmanについては,この業界で知らない人はいないだろう。もともとはスタンフォード大学のArthur Kornberg(1959年ノーベル賞)の弟子で,大腸菌ファージのDNA複製のin vitro系を作る仕事をしていた。現在ダートマス大学で液胞膜融合の研究をしているBill Wicknerと兄弟弟子の関係にある。Randyは20代の若さでカリフォルニア大学バークレイ校にラボを持ち,当時真核生物のモデル系として注目を集め始めていた出芽酵母Saccharomyces cerevisiaeを用いて分泌研究を開始する。生化学者なので,はじめは変異株にも遺伝子にもあまり興味がなかったらしいのだが,言うまでもなく酵母を用いる最大のメリットは遺伝学の強みであり,Peter Novick(現エール大学)という優秀な学生を得て,変異株のスクリーニングに進んでいったのは当然のことであった。
当初23個の相補性群に分類されたsec変異株は,全て温度感受性致死で,高温ではオルガネラ間のさまざまなステップで分泌タンパク質の輸送を止め,増殖も停止する5)。致死変異株が取れるということは,そこに必須遺伝子が関与しているということであるから,つまりオルガネラ間の小胞輸送には非常に重要な分子装置が機能している,という今では当たり前のことを,はじめて明確に示したことになったわけである。
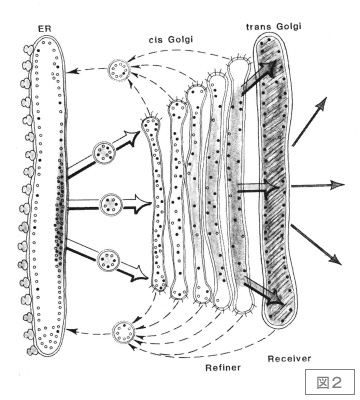 この時代,小胞輸送研究でRandyと並んでひのき舞台に躍り出たのが,当時スタンフォード大学にいたJim Rothmanである。スタンフォードに移る前からMITのHarvey LodishのもとでVSV Gタンパク質を用いた膜タンパク質のアセンブリーの仕事などで名を轟かせていたが,スタンフォードでラボを立ち上げてから,動物細胞の分泌経路に関する生化学的な研究を精力的に開始した。私にとって最も印象的だったのは,1981年にScienceに発表した長いperspective論文6)である。この中でJimは,ゴルジ体はシスとトランスという2つの独立したオルガネラがタンデムに並んだもので,層板構造は一種の蒸留塔のような機能を果たしているのではないかという提案をした(図2)。この考え方は,いずれJim自身は撤回してしまうのだが,私は今でもとても気に入っていて,ゴルジ体の本質はまさにこの蒸留塔の考え方にあるんじゃないかと思っている。Jim Rothmanはその後,VSV Gタンパク質のゴルジ槽間輸送の見事な無細胞系を確立した。これによって,ゴルジ体内輸送が小胞輸送によるという大きなパラダイムを打ち立てたのはよく知られている通りである(時田,中野の節を参照)。
この時代,小胞輸送研究でRandyと並んでひのき舞台に躍り出たのが,当時スタンフォード大学にいたJim Rothmanである。スタンフォードに移る前からMITのHarvey LodishのもとでVSV Gタンパク質を用いた膜タンパク質のアセンブリーの仕事などで名を轟かせていたが,スタンフォードでラボを立ち上げてから,動物細胞の分泌経路に関する生化学的な研究を精力的に開始した。私にとって最も印象的だったのは,1981年にScienceに発表した長いperspective論文6)である。この中でJimは,ゴルジ体はシスとトランスという2つの独立したオルガネラがタンデムに並んだもので,層板構造は一種の蒸留塔のような機能を果たしているのではないかという提案をした(図2)。この考え方は,いずれJim自身は撤回してしまうのだが,私は今でもとても気に入っていて,ゴルジ体の本質はまさにこの蒸留塔の考え方にあるんじゃないかと思っている。Jim Rothmanはその後,VSV Gタンパク質のゴルジ槽間輸送の見事な無細胞系を確立した。これによって,ゴルジ体内輸送が小胞輸送によるという大きなパラダイムを打ち立てたのはよく知られている通りである(時田,中野の節を参照)。
●日米セミナー1981
さて,このRandy Schekman(UCバークレイ)とJim Rothman(スタンフォード)という重要な2人がサンフランシスコ近郊に集まって,酵母の遺伝学と動物細胞の生化学から重要な研究成果を次々に発信し始めたころ,日本学術振興会とアメリカNSFの助成による日米セミナーが京都で行われた(1981年12月)。これは,大村恒雄さん,田代裕さん,故水島昭二さん,そしてDavid Sabatiniが世話人となって,非公開で3日間にわたり行われたもので,私も飛び入りでオブザーバー参加を許され,Günter BlobelとBill Wicknerのバトルなどを目の当たりにすることができた。Jim Rothmanは残念ながら参加していなかったのだが,Randy Schekmanに初めて出会い,講演のときにJim RothmanのこないだのScienceの論文6)をどう思うか,と拙い英語で質問してしまった。そういう個人的なことはあとでゆっくり答えたい,という返事でどっと会場は沸いていたが,私の質問の真意は,酵母でいろいろsec変異株が取れているが,シスからトランスへの輸送の途中で止まるものがどうして取れてこないのだろうというところにあった。その疑問はずっと長いこと棚上げになり,自分自身がその問題を解くことになるなど,そのときには思いもよらないことだった。
●ベイエリアへ
さて,予研の赤松部長からしばらく海外に行ってきていいぞというお許しを得,初めは大村さんや田代さんのお勧めもあってBlobelかSabatiniのところへ留学することを考えていたのだが,結局はRandy Schekmanのところに行くことに決めた。ニューヨークよりもサンフランシスコの方が暮らしやすそうだという安易な理由も若干あったのだが(妻がカリフォルニアがいい!と主張した),やはり何よりも,酵母の遺伝学の強力さに圧倒され,少し勉強してみたいという気持ちになったことが大きかった。私は1983年に,CHO細胞を材料にして,小胞体−ゴルジ体間輸送に損傷をもつ,動物では世界で初めての温度感受性分泌変異株7),というのを単離して得意満面になっていたのだが,その先がとんと進まず,古典遺伝学も分子遺伝学も遺伝子クローニングも自由自在にできる酵母がとても魅力的に見えたのだ。
1981年の日米セミナーでは,Randyは風邪を引いて寝込んでしまい,結局ほとんど話をすることができなかったが,その後手紙をやり取りし,また,たまたま日本細胞生物学会の特別講演に呼ばれたJeff Schatzの代理面接を受けたりして,ポスドクとして受け入れてもらえることになった。1984年から2年間の約束で,家族総勢約5名(5番目はまだ母親のお腹の中だった)がカリフォルニアに渡ったとき,私が31歳,Randyが35歳だった。
ちょうどsec変異株の表現型解析が一段落し,Mitch Bernstein(大学院生)やGreg Payne(ポスドク)といったDNAワークの得意な若手が加わって,いよいよ本格的な遺伝子クローニングを始めようとしていたときであり,そこで新しい分子装置の同定という歴史の1ページに名を連ねることができたのは幸せだったと思う。たまたま選んだSEC12が小胞体の膜タンパク質をコードしており8),また同時にサプレッサーとして取れていた遺伝子がのちに低分子量GTPaseをコードしていると判明する(SAR1)9)など,幸運も重なり,その後の研究人生に限りなくプラスになった留学であったが,同時にそこで得た人のつながりも何ものにも替えがたいものであった。
行ってまもなく,バークレイのSchekman研とスタンフォードのRothman研にUCSFのRegis KellyやPeter Walterのグループなども加わって,タンパク質のトラフィックに関する集まりを定期的に持とうということになった。サンフランシスコ・ベイエリアの関連研究室が親しくつき合う機会が増えて,その縁で私自身がRothman研で実験をさせてもらったこともあった。近い世代でこのころに親しくなり,その後この業界を牽引する友人は数多い。Randy, Jim, Peter Walterもほとんど同じ世代だが,Peter Novick, Susan Ferro-Novick, Scott Emr, Tom Stevens, Greg Payne, François Kepes, Chris Kaiser, Bob Fuller, Bill Balch, Sandy Schmidt, Suzanne Pfeffer, Vivek Malhotra, Paul Melançon, Ben Glickなどなど,ベイエリアから育ったこの人脈が自分の宝物かなとも思っている。
ちょっと脱線するが,この領域からノーベル賞受賞者が何度か出ている。1906年のCamillo Golgiは別格として,先にも述べた1974年のG. Paladeと1999年のG. Blobel,そして1985年のMike BrownとJoe Goldsteinである。実は,1999年のBlobelの受賞のとき,Randy SchekmanとJim Rothmanも共同受賞する可能性が大いにあった。結果的にBlobelの単独受賞となった経緯には,紙面には書けない裏話がたくさんあるのだが,私はやはり小胞輸送研究においてRandyとJimが果たした功績は計り知れないものがあると思う。2002年にこの2人が揃ってラスカー賞を受賞したのは,両方をよく知る1人として,大変うれしいことであった。
●かぐや姫と大発見
さて,2年の留学期間はあっという間に過ぎ,日本に帰る日が近づいていたころ,私はまるで月に帰るかぐや姫のように毎日しくしく泣いて暮らしていた。というのはやや大げさだが,本当に帰りたくなくてたまらない気持ちでいたのである。予研の現職で留学させてもらったので,帰ってちゃんと職があるという実に恵まれた立場だったのに,なぜそんなに悲しんでいたのか。それは,留学中は好きなことをやっていいけど,帰ったらまた動物細胞の仕事に戻ること,酵母は絶対にダメ,と部長に言われていたからである。こんなにおもしろく魅力的な生物を扱うのがこれで終わりなんて悲し過ぎる。帰るぎりぎりまで荷造りもしないで実験室に篭り,あと何かできることはないかと思っていたときに,そうだ,あの遺伝子のシークエンスを決めておこう,と思いついた。sec12温度感受性変異の多コピーサプレッサー遺伝子である。ノックアウトが致死になることは確認していたし,ORFも比較的小さそうである。よし,配列読んでしまえ!と。いまの若い人には想像もつかないでしょう。DNAシークエンスを決めるのは結構大変なことだったのだ。シークエンサーもない。プライマーの合成も簡単じゃない。PCRだってまだなかった。Exo IIIでプラスミドの欠失シリーズを作り,手作業でダイデオキシ反応をやって,手作りのゲルに流してオートラのフィルムを現像し,GATCを自分で書き写していたのですよ。うーむ,化石扱いされそうだ。今なら端を読んでデータベースを調べて終わりだろう。でも,全部読み切るまでどんな遺伝子かわからないドキドキ感がなくなったのは,ある意味つまらなくなったね。
でとにかく,この遺伝子の配列を,帰る直前の1週間で読み切り,日本に持って帰った。間違いはなかったようでORFも読み取れたが,ホモロジー検索をしてもとくに引っかかるものはなく,何をどうすることもできず,しばらく机の前に貼っておいた。さて,部長に言われる通り動物細胞に復帰し,いろいろと新しいテーマも始めたが,どうにも気は晴れない。Schekman研で同僚であったGreg PayneやPeter Böhniが次々にAssistant Professorになってラボを持ち始めたというのに,自分はどうして自分がやりたいことができないんだろうと。まぁ世界中の若い研究者共通の悩みだろうけれど,自分が世界で一番不幸だと思うものである。
さて,そうして悶々として毎日を過ごし,新しいテーマも遅々として進まず,ため息をつきながら机の前に貼ってあるDNA配列を見ていたときに,突然あることに気がついた。出芽酵母は,スプライシングをほとんどしない生物で,イントロンを持つ遺伝子というのはきわめて数が少ない。だから普通ゲノムDNAを読むとそのままORFが取れることになっていて,cDNAを取ったりESTを確認したりする必要がほとんどない。しかし,この遺伝子にはイントロンがあったのだ。GTATGT…TACTAAC…AGというスプライシングのコンセンサス配列を見つけたときは目が点になった。ここを切り出してみると,ORFが上流にさらに延びて190アミノ酸残基の配列が浮かび上がってくる。それをホモロジーサーチしてみると...何と低分子量GTPaseのRasが引っかかってきた9)。
ここから先は夢のような展開であった。こんなおもしろいものを見つけたのだから,一生のお願いだからやらせてくださいと部長に懇願し,絶対に動物細胞に酵母をコンタミさせない,動物細胞の実験もちゃんとやる,などいくつかの約束と引き換えに許してもらった。Secretionに関係したRasホモローグという意味でSar1と名前をつけ(実はRasを裏返しにして命名した),Sec12と同様に小胞体からゴルジ体への輸送に必要であることを証明した9)。東大の安楽泰宏さんに呼んでもらい,小さいながら自分のグループを持つこともできた。Sec12はSar1のグアニンヌクレオチド交換因子であることが,のちにSchekman研のCharlie Barloweによって示され,Sar1のGTPaseサイクルが小胞体からのCOPII小胞形成に必須な役割を果たすことが明らかになっていく。その途中でSchekmanグループとの大変つらく厳しい競争があったりもしたが,岡敏彦君(現九大・分子生命)に始まり佐藤健(K12註)君(現東大・総合文化)に至る精力的な生化学実験のおかげで,いまや究極の完全再構成系が完成して1分子可視化も可能になった(佐藤の節参照)。COPII形成の過程は,小胞輸送のさまざまな素反応の中で最もよく理解が進んでいるステップである10)ということは,おそらくどなたも異論がないだろう。自分がみつけた役者から始まった研究をここまで持ってこられたのは幸せであったと思う。
私のSar1と相前後してPeter NovickがSec4を,Dieter GallwitzがYpt1を同定しており,分泌経路の小胞輸送の過程に複数の低分子量GTPaseが働いているということを世界に先駆けて示すことができたのは,酵母遺伝学の輝かしい成果だった。一方で,GTPaseの分子スイッチとしての概念は,大学院時代にEF-Tuの研究を通じて上代淑人先生に刷り込まれたものだったので,分泌研究へと大きく舵を切り,ずいぶん違う世界を走ってきたと思ったのに,上代先生の手の平からまだ全然抜け出せていないんだな,と孫悟空のような気分にもなったことを覚えている。
●ゴルジ体の問題ふたたび
こんなペースで思い出話を書き連ねていては,どんなに紙面を費やしても足りるものではない。現代に戻るのを急ごう。
Sec12の解析の過程で,これが小胞体とゴルジ体の間をリサイクルしているんじゃないだろうかというのは,バークレイで糖鎖の修飾を調べていて思い始めたことだったが,この発想は,実は先に書いたJim Rothmanの蒸留塔モデルの影響を強く受けている。田代研の山本章嗣さんとJim自身が,シトクロムP450のリサイクルを否定する論文をJCBに並んで発表し,世の中ではかなり風当たりが強い時期だったが,ちょうど同じころ,イギリスMRCのHugh Pelham(彼も私と同い年である)が,KDEL受容体が小胞体とゴルジ体をリサイクルするというモデルを提出し,何となく戦友のような関係になった。Sec12のリサイクルについては,西川周一君(現名大・理)がメカニズムに踏み込み,佐藤健(K10註)君,佐藤美由紀さん(いずれも現群馬大・生調研)の精力的な仕事で見事に証明されただけでなく,膜タンパク質の選別機構というところまで明らかにすることができた11)。
さて,1990年代に入って,いくつかの大きな論争が勃発する。SNAREが膜融合に必要十分なのか(Jim Rothman vs Bill Wickner),細胞分裂の際,ゴルジ体は小さなユニットとして均等に娘細胞に分配されるのか,それとも一旦小胞体に吸収されるのか(Graham Warren vs Jennifer Lippincott-Schwartz)など,大変激しいもので,ゴードン会議で大バトルを何度も目の当たりにした証人としては,いつか書いておきたいという気もする(傍で見ている分にはとてもおもしろかったから)。しかし,何と言っても圧倒的に数多くのグループを巻き込んだ論争は,ゴルジ体内でのタンパク質輸送に関するものだった12)。
これに関してはすでにいろいろなところに書いたし,時田・中野の節でも触れてあるが,簡単に言うと,ゴルジ体を通過していく分泌タンパク質などの積荷が,シスからトランスへとどのように運ばれていくかという問題である。先に少し触れたが,1980年代初頭にJim Rothmanが糖鎖修飾を指標に,VSV Gタンパク質のシスからメディアルへの輸送を試験管内で巧みに再構成し,この過程に小胞輸送が必要であることを示した。それ以前は,シスの槽が次第にトランスへと動いていく槽前進モデルというものも提唱されていたのだが,Jimの仕事以来,ほとんどの教科書が小胞輸送モデル一色になった。ところが,どうもそれでは説明できないという問題がいくつか持ち上がり,また,再構成系でも,小胞で動いているのは積荷ではなくて酵素の方なのではないかという意見が出てきて,世界中のゴルジ研究者を巻き込んだ論争が起こる。小胞輸送を推すのはもちろんJim Rothmanを筆頭として,形態の大御所Lelio Orci,それからGraham Warrenもはじめはどちらかというとこちらの立場であった。一方,そうではないとする側では,酵素がトランスからシスへ移動することによって槽がシスからトランスへ次第に性質を変えていくという,槽成熟モデルが有力になっていった。槽成熟モデルを推した主な人物として,Alberto Luini, Hugh Pelham, Ben Glickなどがいる。
 1998年,Camillo Golgiによるゴルジ体の発見からちょうど100年経った年に,イタリアのパビアで記念シンポジウムが開かれ,世界中から200名以上のゴルジ研究者が集まった。私も招待を受けて参加した(図3)。残念ながらJimとGrahamという大物を欠き,そこでの大バトルは見られなかったが,当然参加者の最大の関心事の1つは,この論争がどのような決着を見るかであった。この少し後だったと思うが,私がイギリスMRCのHugh Pelhamを訪ねたとき,彼のオフィスでディスカッションしている最中に,突然Hughの机上のマックがスリープし,スクリーンセーバーが動き出した。一旦ブラックアウトした後,ブーンと動き出したのはJim Rothmanの写真である。思わず吹き出した。何だいこれは!?と聞くと,ファイトが湧くだろ?との返事だった。1999年,ゴードン会議の夜,HughとJimが立ったまま繰り広げた2時間にわたる激論も忘れられないが,まだ生々しいのでやめておこう。
1998年,Camillo Golgiによるゴルジ体の発見からちょうど100年経った年に,イタリアのパビアで記念シンポジウムが開かれ,世界中から200名以上のゴルジ研究者が集まった。私も招待を受けて参加した(図3)。残念ながらJimとGrahamという大物を欠き,そこでの大バトルは見られなかったが,当然参加者の最大の関心事の1つは,この論争がどのような決着を見るかであった。この少し後だったと思うが,私がイギリスMRCのHugh Pelhamを訪ねたとき,彼のオフィスでディスカッションしている最中に,突然Hughの机上のマックがスリープし,スクリーンセーバーが動き出した。一旦ブラックアウトした後,ブーンと動き出したのはJim Rothmanの写真である。思わず吹き出した。何だいこれは!?と聞くと,ファイトが湧くだろ?との返事だった。1999年,ゴードン会議の夜,HughとJimが立ったまま繰り広げた2時間にわたる激論も忘れられないが,まだ生々しいのでやめておこう。
さて,この話も書き始めるときりがない。1997年に東大から理研に移り,ライブセルイメージングのための高速高感度共焦点顕微鏡の開発を始めていた私は,その開発中の顕微鏡で解く,最初の課題にこの問題を取り上げることに決めた。ポスドクの松浦さんが,今日の細胞とてもよく光っているんですと呼びにきてくれた日,何枚か写真を撮ってみて,これ,色が変わっているんじゃない!?と気がついたときのドキドキは忘れない。私は,槽成熟説の方が有力とは思いつつも,あくまでも中立に両モデルを検証しようと思っていたのだが,見えてきたものは明らかに槽成熟を支持していたからだ。その後,プロトタイプ機で決定的な証拠をいくつか掴み,2005年,勇んでゴードン会議に向かった。途中,ニューヨークのラガーディア空港にいた私に松浦さんが,デコンボリューションによって劇的に分解能が上がったムービーを送信してきてくれたときの興奮,ゴードンで発表後,しばらく拍手が鳴りやまなかったことなど,書きたいことはまだまだあるのだが,本稿ではこのくらいにしておこう。何にしろ,Ben Glickと並んで酵母のゴルジ体が槽成熟するという論文を発表し13,14),大きな論争に決着をつけたと世界中に認めてもらったのは,大変誇らしいことであった。
ただ,1つの問題の解決は,その10倍の新たな問題を提供してくれる。典型的な槽成熟モデルでは,積荷は槽内にじっとしていて,酵素だけがトランスからシスへと逆行輸送すると考えるが,そんな都合のよいことが本当に起こるのだろうか。小胞に乗っているのは酵素だけ,というのは本当だろうか。COPI小胞が重要であるということは私たちも示したが,どうもそれだけとは思えない。1981年にJimの論文を読み,Randyの講演を聞いたときに感じた素朴な疑問に戻り,ゴルジ体内におけるタンパク質の選別輸送を,納得が行くまで理解したいものだと思っている。
●メントラの世界へ
さて,そろそろこのエッセイもまとめに入ろう。この分野,おもしろい問題がたくさんあるのに,わが国では欧米に比べて研究者人口が少なく,これではなかなか太刀打ちできないと考え,裾野を広げて力をつけるべく,「小胞輸送」という特定領域研究を苦労のすえ立ち上げたのは1998年のことであった。その時は,公募なし,計画班員10名のみという小さなグループだったが,4年間で高い評価を得,次の大野さんの特定領域「メンブレントラフィック」につなげることができた。この大野特定は,公募班員を含めると総勢50名以上となり,大勢の研究者が関連の問題に取り組んでくれるようになっただけでなく,一般的にも小胞輸送やメンブレントラフィックという言葉が市民権を得ることに貢献したと思う。とくに広報担当であった吉森さん(大阪大・微研)が,領域ニュースレターやその他の文章の中で,彼一流の軽妙な筆致で「メントラ」を広めてくれた。これからどのような形でこの分野が発展していくものか,まだまだ想像もつかないことがたくさん待っているような気がする。私もこんなエッセイなど書いて懐古している暇があったら,次はどの問題に挑戦しようかと,虎視眈々と獲物を狙わなくてはならない。また新しい話題を作ることをひそかに心に誓い,今日はここで筆を置くことにしよう。
■脚註
私の研究室でこれまでに一緒に仕事をした中で,とくに大きな貢献をしてくれた仲間に2人の佐藤健がいる。1人は,九大の大村研出身で,修士1年から加わってくれた佐藤健君,もう1人は,東工大の吉田研出身で,米国留学後,名大から理研にスタッフとして移ってきてくれた佐藤健君である。この2人,見ての通り漢字で書いても全くの同姓同名であり,しかも理研の私の研究室に,パーマネントの研究員として何年か共に在籍した。理研の人事に,それは困ります,と言われたが,そう言われても本人達も困るだろう。ミドルネームでもつけようか,という提案は2人に却下され,仕方がないので,採用の年度で区別することにした。平成10年採用の佐藤健君をK10,平成12年採用の佐藤健君をK12としたのである。何だかスパイのコードネームみたいで格好いいではないか(と思ったのは私だけか?)。K10は,夫人の美由紀さん(彼女も結婚前から佐藤だった)と一緒に小胞体とゴルジ体のリサイクリングについての仕事をまとめ,米国留学の後,現在は群馬大・生体調節研で線虫のエンドサイトーシスに関する仕事を進めている。一方,K12の方は,私のところに来る前にBill Wicknerのところに留学して,Jim Rothmanとのバトルに関わる仕事をしていたが,うちに来てからはCOPII小胞形成の完全無細胞再構成系を完成し,現在は東大・総合文化で1分子観察へと仕事を発展させている。2人とも,これからこの分野を担う中堅として期待大である。
■文献
1) Milstein, C., Brownlee, G. G., Harrison, T. M., Mathews, M. B.: Nat. New Biol. 239, 117-120 (1972)
2) Blobel, G., Dobberstein, B.: J. Cell Biol. 67, 835-851; ibid. 852-862 (1975)
3) Palade, G.: Science 189, 347-358 (1975)
4) Novick, P. and Schekman, R.: Proc. Natl. Acad. Sci. U. S. A. 76, 1858-1862 (1979)
5) Novick, P., Field, C., Schekman, R.: Cell 21, 205-215 (1980)
6) Rothman, J. E.: Science 11, 1212-1219 (1981)
7) Nakano, A., Nishijima, M., Maeda, M., Akamatsu, Y.: Biochim. Biophys. Acta 845, 324-332 (1985)
8) Nakano, A., Brada, D., and Schekman, R.: J. Cell Biol. 107, 851-863 (1988)
9) Nakano, A. and Muramatsu, M.: J. Cell Biol. 109, 2677-2691 (1989)
10) Sato, K. and Nakano, A.: FEBS Lett. 581, 2076-2082 (2007)
11) Sato, K., Sato, M., Nakano, A.: RIKEN Rev. 41, 45-46 (2004)
12) Rothman, J. E., Pelham, H. R.: Cell 102, 713-719 (2000)
13) Losev, E. Reinke, C. A., Jellen, J., Strongin, D. E., Bevis, B. J., Glick, B. S.: Nature 441, 1002-1006 (2006)
14) Matsuura-Tokita, K., Takeuchi, M., Ichihara, A., Mikuriya, K., Nakano, A.: Nature 441, 1007-1010 (2006)
図の説明
図1.1980年ころの分泌経路のイメージ
J. D. Watson:”Molecular Biology of the Gene” 第3版(1976)より転載。オルガネラ間に明確な境がなく,一旦小胞体に入った分泌タンパク質は,あとは自然に細胞外へ出ていくように見える。バルクフローの考え方に立てばある意味正しいが,オルガネラ間に選別のメカニズムがあるということはほとんど意識されていなかった。第4版(1987)でも同じ図が採用されている。
図2.Rothmanの蒸留塔モデル
小胞体からゴルジ体のシス領域に送られてきたタンパク質は,層板構造の中で仕分けられ,小胞体に戻るべきものは送り返される。唯一トランス槽までたどりついたものだけが分泌顆粒へと進むことができる。この過程を,沸点の違いによって溶媒を分別蒸留する蒸留塔に例えた。文献(6)より転載。
図3.ゴルジ体発見100周年記念シンポジウム
1998年,Camillo Golgiによるゴルジ体の発見の100周年を記念し,Golgiがかつて教鞭を取っていたイタリアPavia大学において開かれた。写真は,Golgi記念館の前に立つCamillo Golgiの彫像と筆者。